
研究内容
私たちは、植物葉緑体における機能制御に興味を持ち、研究を進めています。
植物の大きな特徴のひとつに光合成があります。高等植物の光合成は葉緑体と呼ばれる複数の膜系からなる形態的にも複雑なオルガネラで進行し、CO2固定が行われています。植物にとって光合成は非常に重要な機能であるため、様々な制御機構を備えています。
私たちはその中でも高等植物葉緑体におけるレドックス制御機構について、その生理機能と分子メカニズムの解明を目指し、研究を行っています。
葉緑体のレドックス制御機構では、チオレドキシンと呼ばれるタンパク質がその制御に中心的な役割を果たします。本研究室では、チオレドキシンファミリータンパク質を中心として、以下の具体的な研究項目を設定し、研究を進めています。
1)葉緑体ストロマにおけるチオレドキシンファミリータンパク質の機能制御機構の解明
2)葉緑体チラコイド膜を介した還元力伝達機構の解明
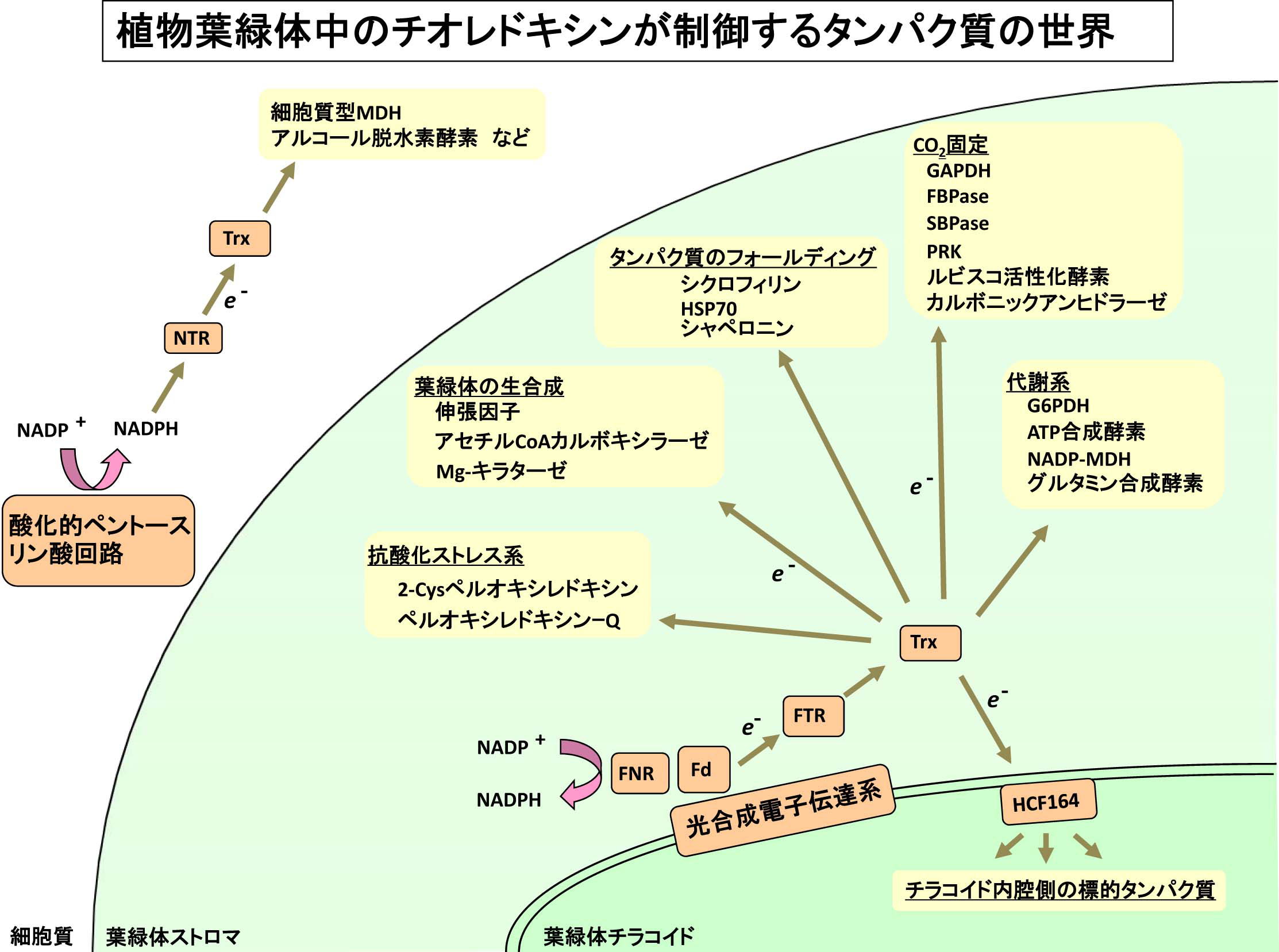 |
【研究例】 「葉緑体m 型チオレドキシン変異体の機能解析」
<背景>
植物の光合成は、葉緑体と呼ばれる細胞内小器官でおこなわれます。光合成の反応に関与する多くのタンパク質は太陽の出ている昼間だけ働くように分子内に制御スイッチを持ちます(図1)。 このスイッチ機構の実体は、タンパク質内にあるジスルフィド結合と呼ばれる化学結合であり、そのスイッチのオン・オフを制御しているのがチオレドキシンと呼ばれるタンパク質です。
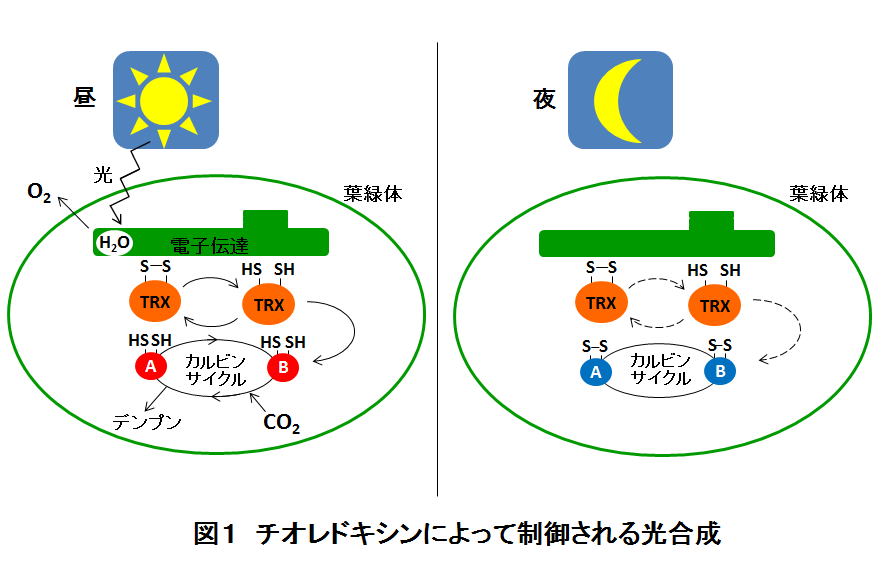
これまで、葉緑体のチオレドキシンは、光合成の際にCO2固定を担うカルビンサイクルの酵素を活性化することが知られていました。また、植物の葉緑体には非常に多くの種類のチオレドキシンが存在することも分かっています。たとえばモデル植物であるシロイヌナズナ(Arabidopsis thaliana)では、5グループ10種類ものチオレドキシンが存在します。しかし、そのメカニズムの研究は、主に試験管内で行われた実験結果を元にしたものであり、実際の植物体内で、数多くのチオレドキシンがどのように酵素の活性化に働いているかは明らかでありませんでした。
<研究概要>
本研究では、シロイヌナズナの突然変異株を用いて、実際の植物体内でのチオレドキシンの生理機能を解析しました。はじめに、シロイヌナズナ葉緑体に存在する5グループのチオレドキシンの存在量を比較すると、m 型チオレドキシンが最も多く存在することがわかりました。そこで、4つあるm 型チオレドキシンのうち、大部分を占めるm 1, m 2, m 4の三重変異株であるtrx m124変異株を取得し、解析を行いました。この変異株では、m 型チオレドキシンの蓄積量が減少しており、その蓄積量にしたがって植物の成長阻害が観察されました(図2)。これまで、m 型以外でのチオレドキシン変異体では成長に影響する表現型は観察されず、m 型チオレドキシンが植物の生育に重要な役割を担っていると考えられます。

三重変異株のうちのひとつは、光合成の電子伝達反応は正常ですが二酸化炭素固定能力が低下していました。この変異体を使って様々な酵素の活性化状態を調べたところ、カルビンサイクル酵素のひとつであるセドヘプツロース-1,7-ビスホスファターゼ(SBPase)の活性化が顕著に抑制されていました。 SBPaseは光合成の律速因子だと考えられており、植物の生育に大きく影響します。これらの結果からtrx m124三重変異株ではカルビンサイクルの活性が抑制されることによって植物の生育阻害が観察されたと考えられます。
これまで、植物葉緑体でのチオレドキシンによる酵素活性制御機構の研究は、試験管内での実験結果をもとに議論されており、SBPaseはm 型ではなく f 型チオレドキシンによって制御されると考えられてきました。また、その他の酵素もf 型によって優先的に制御され、f 型チオレドキシンが酵素活性制御で中心的な役割を担うと信じられてきました。しかし、本研究の植物変異体を用いた解析で、実際の植物体内でSBPaseはm 型チオレドキシンにより制御されていることがわかりました。このことは、in vivoで植物体を用いてこの分野の研究を行うことの重要性を示しています。今後、シロイヌナズナのチオレドキシン変異株をさらに解析することで、チオレドキシンによる新たな制御機構を発見できる可能性があります。
葉緑体内でのチオレドキシンによる光合成の制御システムを明らかにすることで、植物の光合成機能制御の理解を深めることができます。また、将来的には、光合成機能を改変することによって作物の収量を増加させる技術の開発につながる可能性もあります。
.jpg)
