
研究内容
若林研究室では、緑藻クラミドモナスとその近縁藻類を主な材料として、以下の研究に取り組んでいます。
クラミドモナスとは
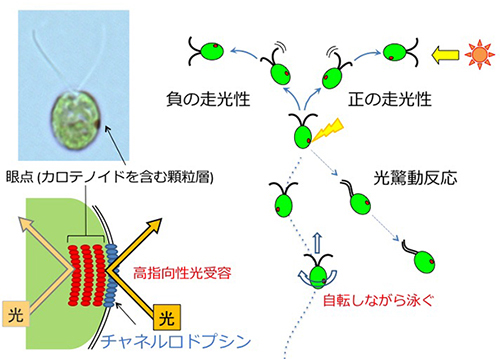
クラミドモナス(学名: Chlamydomonas reinhardtii, 和名: コナミドリムシ)は、淡水に棲む単細胞の緑藻です。長さ約10 μmの2本の繊毛(鞭毛とも呼びますが、最近は繊毛と呼ばれるのが主流になりつつあります)を平泳ぎのように動かして水中を泳ぎます。眼点で光の方向と強度を感じとり、それに応じて遊泳様式を変えます。鞭毛・繊毛運動、光行動反応、光合成、有性生殖、細胞周期など、さまざまな研究分野でモデル生物とみなされています。眼点は赤いカロテノイド顆粒層が光の反射板の役割をし、そのすぐ上の細胞膜に存在する光受容体チャネルロドプシンが光を感じてイオンを透過します。この2つの位置関係で、眼点は指向性の高い光受容を行います。クラミドモナスから見つかったチャネルロドプシンは、現在神経細胞に発現させて光によって神経の興奮を人為的に制御するという「光遺伝学」という分野で大活躍しています。
クラミドモナスの光反応行動の How と Why
クラミドモナスは走光性(光走性)と光驚動反応(光忌避反応)という2種類の光反応行動を示します。走光性とは、2本の繊毛を打つ強さのバランスを調節して、光源方向に泳ぐ(正の走光性)、または光源から遠ざかる方向に泳ぐ(負の走光性)ことです。光驚動反応は、そのとき感じている光よりも極端に強い光を感じたとき、繊毛の波形を一時的に変えて後ろ向きに泳ぐことです。これら2種類の光反応行動は他の多くの運動性藻類にも見られ、クラミドモナスで得られた光反応行動の知見が規範となっています。
走光性の様子。カウンターが5秒になったところで左から、15秒になったところで上から光を当てました。細胞は迅速に遊泳方向を変え、主として正の走光性を示します。
光驚動反応の様子。高速度カメラで撮影し、1/30倍速で再生しています。カウンターが70のところでフラッシュ光を浴びせると、クラミドモナスは「平泳ぎ」型の繊毛運動による前進遊泳を止め、一時的に「ドルフィンキック」型の繊毛運動による後退遊泳をします。
クラミドモナスが走光性の正と負を切り替えるしくみは長らく謎でしたが、私達は以前、細胞内の活性酸素種(ROS)の量の変動がスイッチになっていることを見出しました。一般に光合成生物は、強すぎる光を浴びると光合成の副産物であるROSが大量に発生して死んでしまうため、ROSの発生を抑制するためにあの手この手を使っています。しかし、意外なことに、クラミドモナスは細胞内のROS量が多いときに正の、少ないときに負の走光性を示すことがわかりました(下の写真はペトリ皿にクラミドモナス培養液を入れ、それぞれの薬剤を添加して右から光を当てたもの)。正の走光性を示せば、細胞はより強い光を浴びることになるので、これではまるで自殺行為のように見えます。

私達は、この「クラミドモナスの走光性のROSによる制御」について、まずHow、つまりその分子機構を明らかにしたいと考えています。どのような分子がROS量変動を感知し、どのような分子が繊毛運動調節に関わっているのか。その経路を全て明らかにすることが目標です。そして、Why、つまり走光性がROSによって制御されることが、クラミドモナスが野生で生き抜く上でどのように役立っているのかを知りたいと考えています。生物に物を問えない以上、生物学においてWhy型の質問に回答するのは難しい、あるいは不可能と言っても良いかもしれません。それでも、How質問の答えをたくさん集めて、なんとかしてその答えに近づきたいと考えています。
モデル藻類であるクラミドモナスの行動様式のHowとWhyへの回答は、多様な藻類が関わる有害藻類ブルーム(赤潮など)への対処や、有用物質生産のプラットフォームとして注目を集める藻類の培養法の改善などを通した社会貢献ができると考えています。
繊毛の運動調節メカニズム
真核生物の鞭毛・繊毛は共通して「9+2」という、直径約200 nmの特徴的な内部構造をもっています。9組の2連微小管が2本の中心微小管を取り囲み、各2連微小管の上にはモータータンパク質「ダイニン」が長さ方向に96 nmで結合しています。この精緻な構造は構造生物学者達の心をとらえ、21世紀に入ってからクライオ電子顕微鏡を使った構造解析競争が激しくなりました。9+2構造がなぜ波打ち運動を起こすのか、構造生物学者らの手によって回答されつつあります。
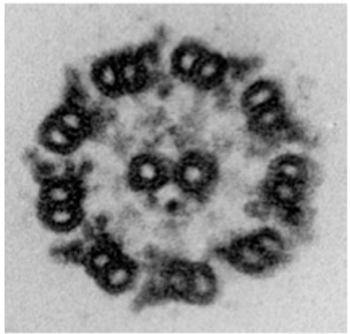
しかし、繊毛はただ同じ運動を繰り返すだけではなく、外部刺激に応じて運動様式を変えます。その分子機構については多くが未解明のままです。クラミドモナスは光受容後1秒にも満たない短い時間に繊毛の運動様式を変えて光反応行動を示すため、繊毛運動調節の良い研究材料です。私達は、クラミドモナスの光反応行動の一番最後のステップである繊毛運動調節の分子機構を明らかにしたいと考えています。
ヒトがさまざまな器官にもつ運動性繊毛の異常により生じる「原発性不動繊毛症候群」と呼ばれる疾患があります。クラミドモナスを用いた繊毛運動調節機構の知見は、こうした疾患の原因やメカニズムの理解に寄与すると考えています。また、技術の分野に「バイオミメティクス(生物模倣)」という言葉があります。よく知られる例として、水をよく弾く蓮の葉の表面構造に似せた撥水フィルムが開発されています。ミクロの世界での鞭毛・繊毛の運動効率は高く、この運動をマイクロマシンの運動制御に取り入れようという考えがあります。私達の研究は、こうした技術にも貢献できると考えています。
これまでの主な研究の日本語解説サイト(過去の所属機関のウェブサイトも含む)
ボルボックスの鞭毛が機能分化していることを発見(2018年)
細胞の運動を「10秒見るだけ」で細胞質ATP濃度がわかる(2021年)
