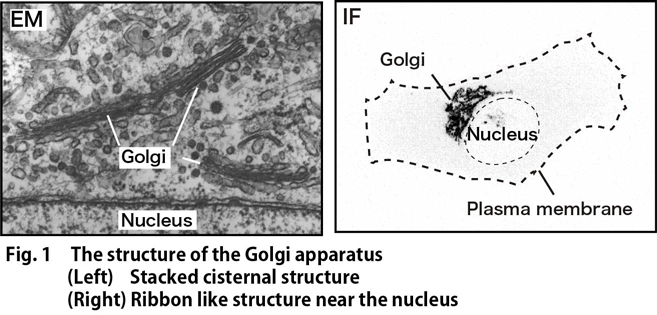
僑儖僕懱偼丆暘斿宱楬偺拞墰偵埵抲偡傞嵶朎彫婍姱偱偁傝丆彫朎懱偱怴婯崌惉偝傟偨僞儞僷僋幙傪庴偗庢傝丆摐嵔傗棸巁婎偺晅壛傗儁僾僠僪嵔偺愗抐側偳偺廋忺傪峴偄丆儕僜僜乕儉傗嵶朎枌側偳偺栚揑抧偵墳偠偰慖暿偟敪憲偡傞婡擻傪扴偭偰偄傞丅僑儖僕懱偺懚嵼偲婡擻偼丆扨嵶朎偺峺曣傗尨惗惗暔偐傜丆懡嵶朎偺怉暔丒摦暔傑偱傎偲傫偳偺恀妀惗暔偵曐懚偝傟偰偄傞丅僑儖僕懱偼丆擷偁傞偄偼憛偲屇偽傟傞儕儞帀幙擇廳憌偱暍傢傟偨戃忬偺峔憿暔偱偁傝丆傎偲傫偳偺愐捙摦暔傗崅摍怉暔偱偼丆滸暯側宍忬偺憛偑愊傒廳側偭偨憌斅峔憿傪庢偭偰偄傞乮Fig.1嵍乯丅偝傜偵愐捙摦暔偱偼丆憌斅偑懁曽偱宷偑傝偁偭偰儕儃儞忬偺崅師峔憿傪宍惉偟偰偄傞乮Fig.1塃乯丅摉尋媶幒偱偼丆偙偺僑儖僕懱偺峔憿宍惉偺暘巕婡峔偲丆峔憿偺惗棟揑堄媊偺棟夝傪栚巜偟偰尋媶傪恑傔偰偄傞丅
僑儖僕懱偼丆暘楐婜偵夝懱偝傟丆柡嵶朎偵嬒摍偵暘攝偝傟偨偺偪偵嵞峔惉偝傟傞丅偙傟傑偱偵変乆偼丆僑儖僕懱偺夝懱暘嶶偑GM130偺儕儞巁壔偵傛偭偰堷偒婲偙偝傟傞偙偲傪柧傜偐偵偟偨乮Nakamura et al., Cell 89 p445, 1997乯丅傑偨堦曽丆僑儖僕懱偺夝懱暘嶶偼暘楐婜偺恑峴偵昁恵偺栶妱傪帩偭偰偄傞偙偲傕柧傜偐偵偟偨乮Yoshimura et al. J. Biol. Chem. 280 p23048, 2005乯丅偝傜偵丆娫婜偺僑儖僕懱偼拞怱懱晅嬤偺旝彫娗偵棈傓傛偆偵偟偰嬊嵼偟偰偍傝丆僑儖僕懱偺嵞峔惉偼丆嵶朎塣摦帪偵恑峴曽岦傪曄壔偝偣傞偨傔偵廳梫偱偁傞偙偲丆傑偨丆GM130偺僷乕僩僫乕僞儞僷僋幙偱偁傞GRSASP65偺儕儞巁壔偑偙偺僑儖僕懱偺嵞峔惉偵廳梫偱偁傞偙偲傕柧傜偐偵偟偰偄傞乮Bisel et. al. J. Cell Biol. 182 p837, 2008乯丅
嵟嬤偺尋媶偐傜丆僑儖僕懱偺峔憿傗婡擻偺晄慡偑傾儈儘僀僪慇堐宍惉傪桿摫偟偰傾儖僣僴僀儅乕昦傗ALS乮嬝堔弅惈懁嶕峝壔徢乯側偳偺恄宱曄惈幘姵傪摫偔壜擻惈傗丆僑儖僕懱偵嬊嵼偡傞僞儞僷僋幙孮偑嵶朎崪奿傗嵶朎嬌惈偺挷愡丆傑偨丆嵶朎撪忣曬揱払宯偺挷愡偵娭梌偟偰偍傝丆偙傟傜偺僞儞僷僋幙偺婡擻晄慡偑丆嵶朎偺娻壔偵娭傢傞偙偲側偳偑師乆偲柧傜偐偵側偭偰偒偨丅偙傟傜偺僑儖僕懱偺峔憿曄壔傗婡擻晄慡偐傜惗偠傞幘姵偼丆乽僑儖僕懱昦乿偲柤偯偗傜傟丆偦偺尋媶偑拲栚傪廤傔偰偄傞乮拞懞挩岹 惗壔妛90 p21, 2018)丅攟梴嵶朎傗僛僽儔僼傿僢僔儏傪梡偄偨尋媶偐傜丆嵶朎儗儀儖丆偦偟偰屄懱儗儀儖偱偺僑儖僕懱傗GM130丆GRASP65, YIPF乮Yip domain family: 屻弎乯側偳偺婡擻傪柧傜偐偵偡傞偙偲偱丆娻傗恄宱曄惈幘姵側偳偺奺庬幘姵偺昦棟偺夝柧傗怴婯帯椕昗揑偺敪尒偑婜懸偝傟傞丅
YIPF僞儞僷僋幙孮偼変乆偑2003擭偵摨掕偟偨僑儖僕懱偵嬊嵼偡傞暋悢夞枌娧捠僞儞僷僋幙孮偱偁傝丆GM130傗GRASP65偲嫤挷偟偰僑儖僕懱傊偺僞儞僷僋幙嬊嵼傗峔憿堐帩偵婡擻偟偰偄傞壜擻惈偑偁傞丅Saccharomyces cerevisiae偵偼丆Yip1p, Yif1p, Yip3p, Yip4p偺係庬偺YIPF偑懚嵼偟丆堦曽丆僸僩YIPF偱偼丆YIPF1~6, YIP1B, YIF1A, YIF1B偺俋庬偑懚嵼偡傞丅傑偨丆Saccaromyces cerevisiae偺Yip1p偍傛傃Yif1p偺儂儌儘僌偑儁傾偲側偭偰暋崌懱傪宍惉偡傞(Yoshida, Y., et al. Exp. Cell Res. 314, 3427亅3443丆2008; Tanimoto, K. et al., Cell Struct. Funct. 36, 171亅185, 2011; Soonthornsit, J., Exp Cell Res, 353, 100-108, 2017乯丅僸僩YIPF偼丆俁庬偺撈棫偟偨暋崌懱1~3傪宍惉偟丆偦傟偧傟僑儖僕懱忋棳乮ERGIC乯丆拞棳乮cis-Golgi乯丆壓棳乮medial-, trans-Golgi, TGN乯偵暘偐傟偰嬊嵼偟偰偄傞乮Fig. 2乯丅
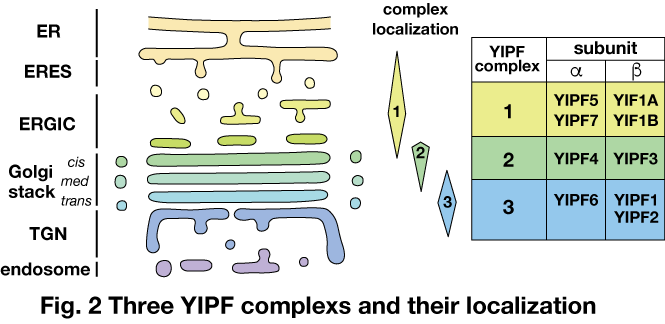
YIPF僞儞僷僋幙偺攝楍傪梡偄偰僞儞僷僋幙攝楍僨乕僞儀乕僗偺専嶕傪峴偭偨偲偙傠丆庒姳偺椺奜傪彍偄偰傎偲傫偳慡偰偺恀妀惗暔偱丆Saccharomyces cerevisiae偺YIPF僞儞僷僋幙4庬偺僆儖僜儘僌慡偰偑懚嵼偡傞偙偲偑妋擣偝傟偨丅傑偨丆摦暔奅乮Metazoa乯傪娷傓Holozoa偱偼丆扨嵶朎惗暔偺Salpingocea rosetta傪娷傔偰傎偲傫偳慡偰偺庬偵丆僸僩YIPF偺偆偪偺6庬乮YIPF1, YIPF3, YIPF4, YIPF5, YIPF6, YIF1A乯偺僆儖僜儘僌偑懚嵼偟偰偄偨丅Holozoa埲奜偺惗暔偱偼丆僸僩YIPF3, YIPF4偵憡摉偡傞僞儞僷僋幙偺懚嵼偼尒傜傟偢丆偙傟傜偺僑儖僕懱拞棳乮cis-Golgi乯偵嬊嵼偡傞YIPF暋崌懱偺抋惗偑丆懡嵶朎壔偲摦暔奅偺恑壔偵廳梫側栶妱傪壥偨偟偨壜擻惈偑峫偊傜傟偨丅傑偨丆峝崪嫑椶偐傜歁擕椶偵帄傞Teleostomi偱偼丆捁椶(Aves)傪彍偄偰丆僸僩摨條偺俋庬偺僆儖僜儘僌偑懚嵼偟偰偄偨丅偙偺偙偲偐傜丆愐捙摦暔偺峝崪壔偵偙傟傜偺儂儌儘僌偺憹壛偑壗傜偐偺栶妱傪壥偨偟偨壜擻惈偑峫偊傜傟偨丅
僛僽儔僼傿僢僔儏偺弶婜敪惗偱偺丆僑儖僕懱偺峔憿曄壔偲偦偺惗棟揑堄媊傪夝愅偡傞栚揑偱僑儖僕懱儅乕僇乕偱偁傞GlcNAcTI-GFP傪敪尰偡傞僩儔儞僗僕僃僯僢僋僛僽儔僼傿僢僔儏傪嶌惉偟偨丅嶌惉偟偨僩儔儞僗僕僃僯僢僋僛僽儔僼傿僢僔儏偺F1悽戙儊僗偵棏妅偺捈宎偑70%掱搙弅彫偟偨棏傪嶻棏偡傞屄懱傪尒弌偟偨丅偙偺彫宆棏妅昞尰宆傪帵偡棏偱偼丆惓忢宆偵斾傋偰嫮偄GFP寀岝偑娤嶡偝傟偨丅僩儔儞僗僕乕儞嶻暔偼GlcNAcTI妶惈傪曐帩偟偰偄傞偲峫偊傜傟傞偨傔丆彫宆棏妅昞尰宆偑僑儖僕懱偱偺GlcNAcTI妶惈忋徃偵傛偭偰婲偙傞壜擻惈偑帵嵈偝傟偨丅GlcNAcTI妶惈忋徃偲偦傟偵敽偆彫宆棏妅昞尰宆偼丆(1) 崅敪尰傪摫偔愼怓懱晹埵傊偺慻傒崬傒丆偁傞偄偼(2)暋悢偺GlcNAcTI-GFP偺慻傒崬傒偵傛偭偰偍偙傞壜擻惈偑峫偊傜傟傞丅偙傟傜偺壜擻惈傪柧傜偐偵偡傞偨傔丆傑偢丆inverse PCR朄偵傛偭偰慻傒崬傒晹埵偺摿掕傪峴偭偨丅
嶐擭傑偱偵丆彫宆棏妅昞尰宆傪帵偝側偄僩儔儞僗僕僃僯僢僋僛僽儔僼傿僢僔儏宯摑偼23斣愼怓懱偵僩儔儞僗僕乕儞偑慻傒崬傑傟偰偄傞偑丆彫宆棏妅昞尰宆傪帵偡屄懱偺僎僲儉偱偼丆23斣愼怓懱埲奜偺晹埵偵僩儔儞僗僕乕儞偺慻傒崬傒偑偁傞偙偲偑柧傜偐偵偟偰偄偨丅崱擭搙偼丆偙偺慻傒崬傒晹埵偺扵嶕傪峴偭偨丅傑偢丆惗柦帒尮娐嫬妛壢偺栘懞嫵庼丒嶁杮彆嫵偲偺嫟摨尋媶偵傛偭偰慡僎僲儉夝愅傪峴偭偨偲偙傠丆25斣愼怓懱偺31514547偐傜31514557偺晹埵乮GRCz11乯傊僩儔儞僗僕乕儞偑慻傒崬傑傟偰偄傞壜擻惈偑柧傜偐偵側偭偨丅PCR偵傛偭偰慻傒崬傒晹埵偺摿掕傪峴偭偨偲偙傠丆幚嵺偵僩儔儞僗僕乕儞偑25愼怓懱偺31514547偐傜31514557偺晹埵乮GRCz11乯傊偺慻傒崬傑傟偰偄傞偙偲偑妋擣偝傟偨丅堦曽丆嬃偄偨偙偲偵丆彫宆棏妅昞尰宆傪帵偡帗傪娷傓siblings偺懡偔偱23斣偍傛傃25斣愼怓懱偺俀僇強傊偺慻傒崬傒偑尒傜傟偨偙偲偐傜丆僎僲儉偺堎側傞晹埵俀僇強傊偺慻傒崬傒偵傛傞僩儔儞僗僕乕儞偺崅敪尰偑彫宆棏妅昞尰宆傪摫偄偨壜擻惈偑帵嵈偝傟偨丅
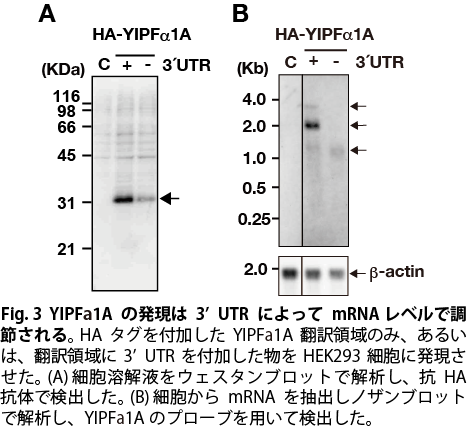
YIPF兛1A偺僐乕僨傿儞僌椞堟偵僱僀僥傿僽偺3乫UTR傪晅壛偟丆HEK293嵶朎偵堦夁惈偵敪尰偝偣傞偲丆僐乕僨傿儞僌椞堟偺傒偲斾傋偰敪尰検偑2攞埲忋崅偔側傞偙偲偑嶐擭搙傑偱偵柧傜偐偲側偭偰偄偨乮Fig. 3A乯丅 僲僓儞僽儘僢僥傿儞僌偱YIPF兛1A偺mRNA偺僒僀僘傪夝愅偟偨偲偙傠丆3乫UTR攝楍偺慡挿偑揮幨偝傟偨偲峫偊傜傟傞栺3300 base偺僶儞僪埲奜偵丆3乫UTR攝楍偺拞墰晅嬤偱揮幨廤寢偟偨偲峫偊傜傟傞栺2000 base偺僶儞僪丆傑偨丆僐乕僨傿儞僌椞堟偺傒偱揮幨廤寢偟偨偲峫偊傜傟傞栺1000 base偺僶儞僪偺3庬偺僶儞僪偑専弌偝傟偨乮Fig. 3B丆栴報乯丅拞偱傕丆栺2000 base偺僶儞僪偺僔僌僫儖偼丆僐儞僩儘乕儖偵斾妑偟偰尠挊偵嫮偔丆mRNA偑憹壛偟偰偄傞偙偲偑帵嵈偝傟偨丅3乫UTR偵傛傞YIPF兛1A偺mRNA偺敪尰検偺曄壔傪掕検揑偵斾妑偡傞偨傔偵丆qPCR偵傛偭偰夝愅偟偨偲偙傠丆僐儞僩儘乕儖偵斾妑偟偰3乫UTR偺晅壛偵傛傝丆mRNA偺敪尰検偑2.4攞偵忋徃偟偰偄偨丅埲忋偺寢壥偐傜丆YIPF兛1A偺敪尰偼丆3乫UTR偺晅壛偵傛偭偰mRNA儗儀儖偱忋徃偡傞偙偲偑柧傜偐偲側偭偨丅
堦曽丆YIPF兛1A偺cDNA ORF偵偼旕嵟揔僐僪儞偑懡梡偝傟偰偄傞偙偲偑敾柧偟偨偨傔丆YIPF兛1A偺cDNA ORF偺僐僪儞傪嵟揔壔偟偰丆YIPF兛1A偺敪尰検偑忋徃偡傞偐偳偆偐傪夝愅偟偨偲偙傠丆梊憐捠傝丆僐僪儞偺嵟揔壔偵傛傝YIPF兛1A偺敪尰検偑尠挊偵忋徃偡傞偙偲偑柧傜偐偲側偭偨乮Fig. 4乯丅 埲忋偺寢壥偐傜丆YIPF兛1A偺敪尰偼丆旕嵟揔僐僪儞偺懡梡偵傛偭偰掅偔梷偊傜傟偰偄傞堦曽偱丆挿嵔偺3乫UTR偺晅壛偵傛偭偰mRNA儗儀儖偱敪尰掅壓偑寉尭偝傟偰偄傞偙偲偑柧傜偐偲側偭偨丅旕嵟揔僐僪儞偺棙梡偵傛傞儕儃僜乕儉偺掆懾偑mRNA偺暘夝傪桿摫偡傞偙偲傗丆挿嵔偺3乫UTR偺晅壛偑mRNA偺埨掕壔傪桿摫偡傞偙偲偑抦傜傟偰偄傞丅偟偨偑偭偰丆YIPF兛1A偺敪尰偼偙傟傜偺挷愡婡峔傪棙梡偟偰岻柇偵挷愡偝傟偰偄傞壜擻惈偑帵嵈偝傟偨丅崱屻丆偙偺挷愡婡擻偺惗棟揑堄媊傪偝傜偵柧傜偐偵偟偰偄偔梊掕偱偁傞丅
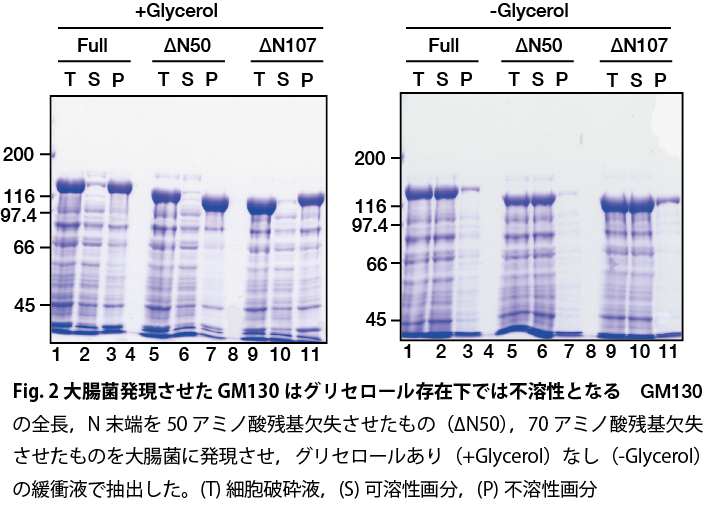
GM130偺峔憿曄壔傪夝愅偡傞夁掱偱丆戝挵嬠偱敪尰偝偣偨GM130僞儞僷僋幙偺惛惢傪峴偭偨丅偟偐偟側偑傜丆偙傟傑偱梡偄偰偄偨拪弌塼乮20mM Hepes-K(pH7.5)丄150mM NaCl丄5% Glycerol乯偱偼丆嵞尰傛偔壜梟壔偱偒側偄偙偲偑柧傜偐偲側偭偨乮Fig.2塃恾乯丅慜擟幰偺忦審傪惛嵏偟丆攟梴忦審傪専摙偟偨偑丆偙傟傑偱偺忦審偱偼廫暘偵壜梟壔偱偒側偐偭偨丅偦偙偱丆拪弌塼偺慻惉偺嵞専摙傪峴偭偨丅偦偺寢壥丆僞儞僷僋幙埨掕壔嵻偲偟偰揧壛偟偰偄偨僌儕僙儘乕儖傪拪弌娚徴塼偐傜彍嫀偡傞偲丆GM130偑尠挊偵壜梟壔偡傞偙偲偑柧傜偐偲側偭偨乮Fig.2嵍恾乯丅崱屻偼丆壜梟壔偟偨GM130傪梡偄偰丆GM130奺庬寢崌僞儞僷僋幙傗GM130儕儞巁壔偵傛傞GM130暘巕廤崌忬懺偺曄壔傪夝愅偡傞丅
YIPF偺兛僒僽儐僯僢僩兝僒僽儐僯僢僩偼俀検懱傪宍惉偡傞偑丆兛僒僽儐僯僢僩偲兝僒僽儐僯僢僩傪摍検敪尰偝偣側偄偲丆僑儖僕懱偺惓偟偄晹埵偵攝憲偝傟側偄壜擻惈偑偁傞丅堦曽丆堚揱巕摫擖偵傛傞堦夁惈敪尰偱偼丆YIPF兛僒僽儐僯僢僩偺敪尰偑YIPF兝僒僽儐僯僢僩偵斾傋偰戝曄掅偔乮師崁嶲徠乯奜棃惈堚揱巕摫擖偵傛傞夝愅偑崲擄偱偁偭偨丅偙傟傪崕暈偡傞偨傔丆YIPF偺兛僒僽儐僯僢僩偲兝僒僽儐僯僢僩傪摍検敪尰偡傞偨傔偺僔僗僥儉偺峔抸傪峴偭偨丅 僂僀儖僗桼棃偺P2A攝楍偼丆攝楍捈屻偱東栿傪廔椆偝偣傞偑丆P2A偵懕偔摨偠僼儗乕儉偺東栿椞堟傪堷偒懕偒東栿偝偣傞惈幙傪帩偭偰偄傞丅偙偺惈幙傪棙梡偟偰丆俀庬偺僞儞僷僋幙傪堦偮偺mRNA偐傜崌惉偡傞偙偲偑偱偒傞乮Felipe, P. et al. Trends Biotechnol 24, 68亅75, 2006乯丅YIPF兝僒僽儐僯僢僩偺東栿椞堟偺屻晹偵P2A傪夘偟偰兛僒僽儐僯僢僩偺東栿椞堟傪攝抲偡傞偙偲偱丆YIPF偺兛僒僽儐僯僢僩偲兝僒僽儐僯僢僩傪摍検敪尰偡傞敪尰僾儔僗儈僪傪嶌惉偟偨丅梊旛揑幚尡偐傜丆YIPF偺兛僒僽儐僯僢僩偲兝僒僽儐僯僢僩偑摨摍偺検敪尰偱偒傞偙偲偑妋擣偝傟偨丅崱屻偼丆僔僗僥儉傪嵟揔壔偟偰丆YIPF偺嬊嵼壔婡峔偺夝愅偵梡偄傞丅
YIPFa1A偺敪尰偑僐僪儞偺嵟揔壔丆傑偨丆3'UTR偺晅壛偵傛傝憹壛偟偨偙偲偐傜丆椉幰傪慻傒崌傢偣偰偝傜偵敪尰忋徃偑尒傜傟傞偐偳偆偐専徹傪峴偭偨丅 YIPF兛1A偺mRNA検偼丆東栿椞堟偺僐僪儞嵟揔壔媦傃3'UTR偺晅壛偺偄偢傟偵傛偭偰傕憹壛偟丆傑偨丆僐僪儞嵟揔壔偟偨東栿椞堟偵3'UTR傪晅壛偡傞偲丆憡壛揑偵偝傜偵mRNA検偼憹壛偟偨丅堦曽丆嬃偔傋偒偙偲偵丆嵟揔壔偝傟偨YIPFa1A偺東栿椞堟偵YIPFa1A偺3'UTR傪晅壛偡傞偲丆梊憐偵斀偟偰丆僞儞僷僋幙敪尰検偑尠挊偵掅壓偡傞偙偲偑柧傜偐偲側偭偨乮Fig. 3乯丅偙偺寢壥偼丆YIPFa1A偺3'UTR偑愭峴偡傞東栿椞堟偺東栿岠棪偵懳墳偟偰僞儞僷僋幙敪尰検傪挷愡偡傞婡擻傪帩偮偙偲傪帵嵈偟偰偄傞丅崱屻偼偙偺挷愡婡峔偺夝愅傪峴偆丅
